
[Cap au seuil|Sommaire rapide] Avant propos | espace | la vie | monde | les écologies | cénologie | sciences naturelles | interlude | protection de la nature | arbres | anthropocène | catastrophe | herbier/coquillier | ouverture : régions naturelles
Les deux (ou trois) écologies < Pour la cénologie > Sciences naturelles = sciences humaines
Texte publié le 29 décembre 2020 et mis à jour pour la dernière fois le 14 février 2025 (35 révisions).
Ces catégories ambiguës, superfétatoires, déficientes rappellent celles que le docteur Franz Kuhn attribue à certaine encyclopédie chinoise intitulée Le marché céleste des connaissances bénévoles. Dans les pages lointaines de ce livre, il est écrit que les animaux se divisent en a) appartenant à l’Empereur, b) embaumés, c) apprivoisés, d) cochons de lait, e) sirènes, f) fabuleux, g) chiens en liberté, h) inclus dans la présente classification, i) qui s’agitent comme des fous, j) innombrables, k) dessinés avec un très fin pinceau de poils de chameau, l) et caetera, m) qui viennent de casser la cruche, n) qui de loin semblent des mouches.
Jorge Luis Borges
Aujourd’hui dominante dans les domaines de la description et de la classification des milieux naturels[1]Parts de ce texte ont paru sous le titre « What has become of our cenosis? » dans Plant Sociology n°58/2, 2021, et d’autres sous le titre « Éléments pour la description des cénoses … Continue reading, la phytosociologie est l’héritière d’une longue tradition remontant aux balbutiements de l’écologie, avec la phytogéographie. Elle s’intéresse aux groupements de plantes en un lieu donné, et se donne pour tâche de décrire les « associations », lesquelles sont définies en 1910 par Charles Flahault et Carl Schröter : « groupement végétal de composition floristique déterminé, présentant une physionomie uniforme, croissant dans des conditions stationnelles uniformes[2]C. Flahault , C. Schröter , Rapport sur la nomenclature phytogéographique, in IIIe Congrès international de botanique, Wildemann, Bruxelles, 1910, Actes du IIIe Congrès international de … Continue reading », définition que complètent dès 1913 Josias Braun et Ernst Furrer, « … et possédant une ou plusieurs espèces caractéristiques[3]Josias Braun et Ernst Furrer, « Remarques sur l’étude des groupements de plantes », in Bulletin de la Société Languedocienne de Géographie n°36, 1913, 20-41. ».
En insistant sur le regroupement d’espèces, plutôt que sur les grands ensembles dynamiques, la phytosociologie prend son envol et s’autonomise rapidement du « courant écosystémique », notamment sous l’impulsion de la Station Internationale de Géobotanique Méditerranéenne Alpine où œuvre le jeune Josias Braun(-Blanquet), originaire de Zürich (d’où le nom d’école zuricho-montpelliéraine). Malgré l’existence, voire la persistance d’écoles alors concurrentes, comme, dans le Nord de l’Europe, l’école d’Upsal, développée à partir du même fonds phytogéographique, sous l’impulsion de chercheurs, tels que Robert Fries ou Gustav E. du Rietz, qui favorisent une vision plus physionomiste, indépendante de la floristique stricte, et par conséquent plus en phase avec le « courant écosystémique » (et chez qui elle trouvera un accueil plus favorable) d’outre-Atlantique, elle s’est finalement imposée jusque dans les instances européennes, notamment par le biais du programme Natura 2000.
De la cénologie
Mais la phytosociologie ne représente, en vérité qu’une partie d’un ensemble épistémologique plus vaste, et qu’on a déjà nommé ici biocénologie (ou cénologie, ou cénotique, parfois même l’écobiocénologie, et encore Synécologie). La phytosociologie est en fait une phytocénologie, et s’intéresse par conséquent aux cénoses végétales. Les cénoses animales, les cénoses fongiques, les cénoses bactériennes pourraient ou devraient, en toute logique, former les autres parties de la cénologie, avec la zoocénologie, la mycocénologie, etc.
Pourtant, chose conceptuellement frappante, il n’y a pas, ou pratiquement pas, d’études dédiées aux bactéries, très peu aux groupements mycologiques [4]Pour ces deux domaines de recherche, toutefois, il faut préciser leur singularité biologique ou leur originalité épistémologique. En effet, dans les deux cas, on constate une très forte … Continue reading, et encore moins aux groupements animaux. C’est un mystère lié à l’histoire des sciences, et probablement aussi à l’histoire tout court, dont l’explication, que j’ai recherchée par ailleurs, devrait nous orienter sur le cours pris par l’écologie depuis les années 60-70.
Si les apports de la phytosociologie n’ont commencé à infiltrer la zoologie que dans les années 1950, les questions relatives aux communautés animales, et aux liens qu’elles entretiennent avec les communautés végétales ou les écosystèmes, sont évidemment beaucoup plus anciennes. La biocénotique est un sujet en pleine effervescence dans ces mêmes années. Un congrès historique, le premier strictement dédié à l’écologie qui se déroule en Europe, se tient à Paris, sous l’égide du CNRS, en 1950, qui fait le point sur la question, associant botanistes et zoologues[5]Actes du colloque sur l’écologie de 1950 à Paris, in Colloques Internationaux du Centre National de la Recherche Scientifique, n°XXXIII, Paris, C.N.R.S [Note de Pascal Acot : ‘Les Actes du … Continue reading. Plusieurs chercheurs ont proposé (ou ont récemment proposé, alors) des synthèses théoriques tout à fait opératoires ; comme l’indique le célèbre article de Quézel & Verdier de 1953, la question est clairement de savoir si les méthodes de la phytosociologie sont applicables à l’étude des groupes d’animaux[6]Pierre Quézel et Paul Verdier, « Les méthodes de la phytosociologie sont-elles applicables a l’étude des groupements animaux ? Quelques associations ripicoles de carabiques dans le Midi de la … Continue reading. De nombreux chercheurs ont identifié des zoocénoses[7]On pourrait citer Rabeler, Künhelt, Strenzke, Sacchi, Schwenke et Quézel & Verdier, Amiet, Mouthon, Frank, etc. tout ceci largement concentré dans les années 50.. Il devient alors possible d’imaginer le synopsis d’un prodrome de zoocénose, au moins au niveau d’un clade : Orthoptères, Coléoptères, Gastéropodes…
Par la suite, dans les années 60 à 80, on observe le développement des études d’assemblages d’espèces, qui sont aujourd’hui classiques : les biocénoses littorales associant algues et animaux, étudiées par Molinier ou Boudouresque[8]Roger Molinier, « L’étude des biocénoses marines du Cap Corse », in Vegetatio n°9, p.121–192 & 217–312, 1960 ; Charles-François Boudouresque, « Recherches sur les concepts de … Continue reading, la bionomie de Pérès & Picard[9]Jean-Marie Pérès & Jean Picard, « Nouveau manuel de bionomie benthique de la mer Méditerranée », Recueil des Travaux de la Station Maritime d’Endoume n°47(31), p.1-137., qui servira bientôt de base aux différents référentiels d’habitats marins. Les communautés du sol sont également bien étudiées, par exemple les assemblages de collemboles ou de vers de terre, ou l’engouement plus récent pour la rhizosphère. Enfin, les fameuses typologies de rivières, instituées par Huet, complétées par Illiès & Botosaneanu, puis Verneaux[10]Marcel Huet, « Aperçu des relations entre la pente et les populations piscicoles des eaux courantes », in Schweizerische Zeitschrift für Hydrologie n°11, p.332–351, 1949 ; Jean Illiès … Continue reading, représentent un dépassement des simples préoccupations cénotiques, puisqu’elles s’attachent davantage à la notion de population ou de peuplement, plutôt qu’aux véritables communautés écologiques, en l’occurrence celles des poissons, ou celles des larves aquatiques d’insectes.
Plusieurs synthèses ont été publiées dans les années 1960 et 1970. En 1962, l’Américain Robert Whittaker a réalisé une synthèse impressionnante sur la question des communautés biologiques avec une énorme bibliographie[11]Robert Whittaker, « Classification of Natural Communities » in The Botanical Review, n°28/1 : pour les cénoses animales, voir p.62-69 ; dans la bibliographie tous les auteurs cités jusqu’en … Continue reading. Les actes d’un autre congrès historique, qui s’est tenu en 1960, ont été publiés en 1966 sous la direction de Reinhold Tüxen : Biosoziologie[12]Reinhard Tüxen (éd.), Biosoziologie, La Hague, Junk, 1966.. Enfin est publié l’un des rares manuels sur le sujet[13]Roger Molinier & Pierre Vigne, Ecologie et biocénotique, Delachaux & Niestlé, 1971..
Dans les années 80 et 90, cependant, le fil s’amincit progressivement ; quelques auteurs allemands poursuivent le travail de leurs prédécesseurs. L’un des plus remarquables, et aussi l’un des derniers, est le botaniste Harro Passarge (sur les oiseaux), mais aujourd’hui, hormis quelques travaux isolés, les cénoses ne sont plus considérées.
La cénose et autres types de groupements
Avant toute chose il nous faut comprendre ce qu’est le fondement de la cénologie, à savoir la cénose.
La phytosociologie, on l’a dit, recense les associations, qui sont des groupements écologiques. La question de savoir si ces associations sont de véritables cénoses ou une partie de cénoses plus vastes n’est pas toujours réglée, mais pour l’instant elle ne nous intéresse pas. Nous la retrouverons bientôt. Disons que les cénoses de la phytosociologie, pour des motifs historiques, sont les associations. Ces associations, nombreuses et variées, peuvent être regroupées en ensembles plus vastes, et c’est ainsi que la cénologie végétale se structure en un système intégratif structuré en quatre rangs, un peu à la manière de la systématique classique (où l’espèce serait figurée par la synespèce, ou groupe d’espèces de rang inférieur, l’association). Ces rangs sont donc l’association, puis l’alliance, l’ordre, la classe. Avec le temps (une centaine d’années), il a été jugé parfois utile de recourir, comme dans la systématique classique, à des paliers intermédiaires : sous-classes, sous-alliances, etc. Ce qu’il faut tenir à l’esprit, c’est que, même structurés en système hiérarchique, ces groupements sont des cénoses : la classe est une cénose, l’ordre est une cénose, l’alliance est une cénose, l’association est une cénose.
Les cénoses se distinguent d’autres types de regroupements d’individus ou d’espèces différentes. Chacun sait qu’il est possible d’opérer n’importe quel regroupement d’éléments partageant un trait commun (comme l’évoque Borges en exergue). Ces groupes peuvent être écologiques, ou non : les plantes comestibles et les plantes non comestibles par exemple ; les animaux noirs et blancs ; les animaux qui appartiennent à l’Empereur ou ceux qui ont cassé la cruche. C’est d’ailleurs le travail, en dynamique perpétuelle, de la taxonomie et de la systématique. On sait également que certains groupements concernent plus particulièrement la biologie, et peu ou prou à l’écologie. Qui ne connaît pas la différence entre vivaces et annuelles par exemple, ou encore entre herbivores et carnivores ? Ou bien encore les membres d’une classe d’âge ?
Clades, communautés, assemblages, populations, guildes… tous ces termes existent, sont utilisés et nous sont à peu près familiers : chacun avec ses particularités, ses définitions ou ses compositions particulières.
Avant toute chose, séparons bien les domaines d’intervention : certains groupements relèvent de la génétique ou de la systématique, d’autres de comparaisons et de ressemblances en termes biologiques ou physiologiques, et d’autres encore sont strictement écologiques (au sens de strictement écobiocénotiques).
J’appelle ici communauté (ou communauté biotique), ces groupements d’éléments disons biologiques, sans distinction qualitative particulière.
De quelle manière un organisme peut-il être lié à une communauté ?
- Par l’espace et le temps (n’oublions pas le temps, soyons lamarckiens !), c’est-à-dire les deux catégories a priori non pas de la sensibilité au sens kantien, mais bel et bien de l’être intime au sens bergsonien ou de l’être au monde au sens heideggerien.
- par sa famille : il est l’enfant, le descendant et, potentiellement, le parent, l’ascendant ;
- par l’espèce et le genre (au sens sexuel) : il est chien ou chat, et il est mâle ou femelle ;
- par la biologie : celle liée aux éléments précédents, qui sont ceux disons intrinsèques à l’espèce (ceux du dedans, et en particulier la sexualité/reproduction), mais aussi ceux liés aux dehors, à savoir le métabolisme : la respiration/nutrition. En ce sens il est donc lié par une relation d’obligeance à cette ressource ;
- enfin, même si cela ne concerne que l’homme, il est lié à la noosphère, au langage, à la technique et à la culture, au monde symbolique, au sacré, à l’art et à la mort.
En somme, six dimensions possibles s’accordent pour former une communauté possible.
Mais en poussant plus loin notre raisonnement, il apparaît comme une évidence que la dernière dimension évoquée ne concerne pas d’autres organismes que l’espèce humaine, nous pouvons le laisser de côté (je rappelle que l’écologie s’intéresse à la biosphère ; la noosphère est un domaine de recherche encore différent) : il reste cinq dimensions.
Ensuite, la distinction des domaines relevant de la famille et de l’espèce paraissent, en réalité, ressortir de la même dimension, celle qui touche à la sexualité et à la procréation, c’est-à-dire à la fois paradoxalement à l’individu comme à l’espèce. On peut donc les associer : restent quatre dimensions.
Cette dernière remarque est valable également pour la distinction des domaines du temps et de l’espace, puisqu’il est impossible dans notre univers, de n’être pas à la fois hic et nunc. On peut donc en conclure que trois dimensions dominent, et qui relèvent donc de la spatio-temporalité, de la phylogénie, et enfin par la satisfaction des besoins vitaux (hors sexualité), physiologiques et biologiques, et donc aux ressources (essentiellement la trophie, c’est-à-dire le mode de nutrition).
On aurait donc trois grands types de regroupements possibles : A : la dimension génétique (mais qui associe l’individu à sa famille, et son espèce à son clade !) ; B : la dimension biologique (et en particulier la nutrition) ; C : la dimension écologique.
Les termes de la littérature sont aujourd’hui extrêmement flous et se substituent les uns aux autres allégrement. J’arrête ici les définitions qui suivent : les individus projetés dans la première dimension forment le clade ; dans la deuxième dimension l’assemblage ; dans la troisième dimension le type biologique. Si l’on considère cette fois en même temps : des individus d’un clade et d’une même dimension spatio-temporelle A+B forment une population ; les individus de même coordonnées spatio-temporelles attachés aux mêmes ressources B+C des guildes ; les individus de même clade attachés aux mêmes ressources A+C un peuplement. Si l’on considère cette fois les trois dimensions contemporainement, A+B+C, le terme approprié est précisément celui de cénose.
Implications conceptuelles et épistémologiques
La communauté (ou communauté biotique) est un groupement d’individus biologiques, sans spécification, c’est-à-dire neutre.
On a dit que l’être vivant était un être de la relation. Ces relations peuvent être de trois ordres, trois dimensions ontologiques : phylogénétique (phylogénie), géographique (choro- et phénologie = territoire ou choréologie) et ressourcielle (biologie) : soit, à bien y regarder, les trois piliers de l’écologie : ce qui croise, au passage, le découpage par ailleurs classique de l’écologie en synécologie (les espèces), démécologie (les populations), et autoécologie (les individus[14]En réalité, c’est plus compliqué, comme d’habitude. Rioux distingue, de son côté, trois types de groupements : la cénose, la société et la foule. La foule est un groupement sans autre … Continue reading).
Épistémologiquement, on pourrait résumer cette tripartition de la manière suivante. Les liens phylogénétiques regroupent deux types de relations ; celles liées d’une part par le champ de relation de l’individu avec ses proches, 1. sa « famille », puis avec ses ancêtres et descendants plus ou moins lointains, 2. son espèce, puis son genre, et ainsi de suite ; cette relation est inconsciente dans le second cas ; elle est au contraire éminemment structurante dans le premier dès lors qu’ils sont motivés par des relations familiales, c’est-à-dire génétiques, on peut évoquer un domaine familial ou écogén(et)ique (οικογενειακός). Lorsque les liens sont simplement ceux de la coordonnée spatio-temporelle, ils semblent « hasardeux » ; en réalité cette dimension choréophénologique est pratiquement fondamentale, ontologique, fondatrice même de l’être, en tant tout d’abord qu’individu, avant même son appartenance à une famille ou à une espèce. Enfin, dans le domaine plus précisément lié aux ressources, c’est un lien intéressé, certes, donc plus ou moins indépendant de toute destinée ou de hasard, et qui relève des choix individuels, familiaux ou populationnels ; mais ceci est vrai jusqu’à un certain point, car que dire des communautés d’algues ou de champignons, de lichens, ou même de cnidaires ou de poissons ?
On a beau retourner le problème dans tous les sens, on ne voit pas comment toute communauté pourrait s’abstraire, à volonté, de l’une des trois dimensions, si l’on veut bien que trois dimensions structurent les groupements.
C’est précisément la nature du milieu (Ungebung ou fûdo) que de synthétiser en un seul terme ce que j’allais appeler l’appétence (Deleuze dirait l’aguet) du vivant à la relation.
Le philosophe Lockwood a bien montré, d’ailleurs, toutes les impasses conceptuelles auxquelles ces différents types de regroupements peuvent aboutir[15]Jeffrey A. Lockwood, « The Ontology of Biological Groups : Do Grasshoppers Form Assemblages, Communities, Guilds, Populations, or Something Else ? » in Psyche : A Journal of Entomology, en ligne : … Continue reading. En se penchant sur le cas des groupements des criquets dans la littérature, il a montré combien étaient confuses ces idées pour le chercheur, le scientifique, de surcroît, qui souvent manque de rigueur conceptuelle (un comble).
En vérité, tout groupement biologique est une cénose : toute communauté est une cénose. Certaines sont étudiées d’un certain point de vue (et on les appelle population, guilde, peuplement, ou bien assemblage, type biologique, clade), mais ce ne sont jamais que des cas particuliers de la cénose, qui, s’ils ont un aspect heuristique ou technique commode, n’en demeurent pas moins des constructions de l’esprit.
Seule la cénose existe, si l’on veut, indépendamment de toute syntaxonomie possible ou préalable. Elle est en fait la communauté : « every commmunity beings irrespective of its nature[16]Mauritz Franz Mörzer Bruyns, On biotic communities [1947], Station Internationale Géobotanique Méditerranéenne et Alpine, 1950.».
Notre démonstration par l’absurde n’a finalement rien démontré, mais au moins a-t-elle le mérite de souligner le vrai cœur du problème : si la cénose est fondamentale, comment se fait-il que les écologues aient, peu à peu, délaissé les termes de cénologie, de cénotique et de cénoses ? Est-ce simplement du fait de la confusion entre notions à peu près équivalentes qu’ils ont été progressivement marginalisés ? Probablement pas, puisque les autres restent, au contraire, bien vivants (même si souvent confondus).
Je ne pense pas qu’on puisse supposer, à partir de la simple lecture des articles ou des ouvrages qui parlent des écosystèmes, d’une quelconque faille conceptuelle : comme on l’a vu la cénose est le terme le mieux adapté (et celui qui, sous l’avatar d’association phytosociologique a eu le plus de succès).
Je suppose en conséquence qu’il y a une raison idéologique, qu’accompagnent en sus des raisons purement techniques. Par exemple le développement de la statistique dans les études d’écologie a poussé les « faunistes » , qui n’ont généralement pas adopté le modèle de Braun-Blanquet, à quitter la cénologie, qui porte une part non négligeable d’interprétation subjective, au nom d’une prétendue objectivité numérique[17]Gisin (1964) se méfiait des diagrammes en treillis qui selon lui étaient des non-sens ; Gounot lui-même (1961) mettait en garde à l’absence de recul devant les équations ; Guinochet (1968) : … Continue reading. Les termes et concepts de facteurs, gradients, traits, tous mesurables (quantitatifs), devenus prépondérants, ont éclipsé les fidèles, les compagnes, les caractéristiques, toutes interprétables (qualitatifs)… C’est un fait. C’est donc je pense sous couvert d’un scientisme qui ne dit pas son nom que la cénologie a été abandonnée (ou laissée à des botanistes, ce qui revient à peu près au même étant donné le champ de la formation botanique actuel, en France tout du moins).
Il s’ensuit qu’en toute logique la cénose apparaît bel et bien comme la véritable unité minimale de l’écologie (plutôt que l’écosystème qui, en quelque sorte, n’en est que l’extension maximale, à la condition qu’on l’appréhende comme biome, un bioèce avec un biocène).
Pour une cénologie renouvelée
Philippe Julve a pour habitude de dire à ses interlocuteurs que la phytosociologie est comme la musique de Bach ou de Mozart : elle ne sert à rien. C’est une très belle non-définition, parce qu’elle insiste non seulement sur le caractère arbitraire (c’est-à-dire langagier, c’est-à-dire humain) de la phytosociologie comme classification, et comme toute classification ; mais elle souligne de surcroît la part subjective de la pratique, le côté assimilable à un savoir-faire ou à un talent, en tout cas à une création personnelle, qui sera, ou non, partagée par des pairs, à défaut d’être mathématiquement prouvée.

Définition
La biocénotique (ou biosociologie, ou synécologie) cherche à produire une classification organisée (un synsystème) des groupements d’êtres vivants (cénoses), à partir de leur composition spécifique récurrente, à l’échelle préférentielle du clade. Ainsi on pourra parler de mycocénologie, de zoocénologie, mais également de malacocénologie, d’arthropodocénologie, mais encore d’hexapodocénologie ou d’arachnidocénologie, etc.
Épistémologie
1. La biocénotique est une branche de l’écologie. Comme le souligne justement Rioux, elle est la branche qui précisément s’intéresse aux êtres vivants et à leurs relations (il parle alors d’écologie éthologique) ; elle s’oppose si l’on veut à l’écologie mésologique, qui étudie plutôt les flux de matière et d’énergie.
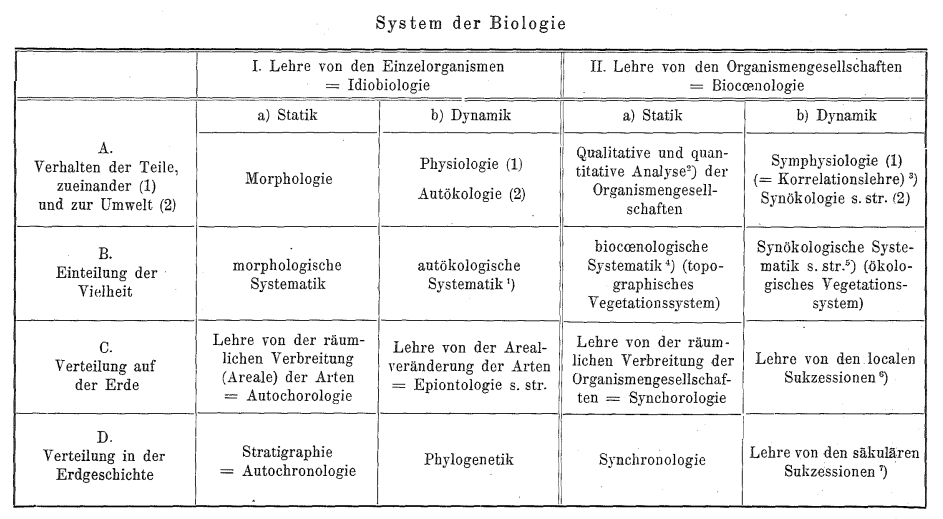
2 L’écologie intègre, selon Gams[18]Helmut Gams, « Prinzipienfragen der Vegetationsforschung. Ein Beitrag zur Begriffsklärung und Methodik der Biocoenologie » , loc.cit., le système de la biologie : selon ce dernier, les domaines de la biologie sont au nombre de quatre : la relation entre espèces et entre espèce et environnement, la classification, la répartition, l’histoire ; ces domaines sont considérés soit du point de vue statique, soit d’un point de vue dynamique.
3 Elle s’appose à la biogéographie qui décrit les grandes formations ou les grands biomes. Il y a un point de rencontre (qui est un point de bascule) entre les deux et qui concerne généralement ce que nombre d’écologues ou de botanistes appellent la formation, même si ce terme est fortement discuté dès son apparition chez Humboldt. Lorsque les cénoses sont devenus assez grandes et que des facteurs abiotiques « prennent le dessus » (climat, sol, roche), il semble que l’on passe dans une autre dimension, qui est celle du paysage (par le moyen du saut quantique).
4 Elle est donc aussi une synthèse[19]Gams, loc.cit., p.298 ; Rioux, op.cit., p.129 ; Mörzer Bruyns, op.cit., p.1-3. des connaissances des cénoses selon leurs traits singuliers, c’est-à-dire dans les domaines de la systématique, de la génétique (ou la phylogénie), de la choréo(chrono)logie, de l’éthologie, de la physiologie), de la morphologie.
Méthode
Sa méthode est descriptive à la fois inductive et inductive — abductive (voir le chapitre suivant) ; elle se fonde sur les sciences naturelles, la biologie et la génétique, ainsi que sur la taxonomie et la cladistique. La méthode peut se décomposer en plusieurs temps, selon le degré de précision qui intéresse le chercheur. Celle-ci, dûment et régulièrement éprouvée par les phytosociologues, suppose des phases de terrain et de laboratoire, ainsi que des phases d’analyse et de synthèse, mais aussi des phases objectives et des phases subjectives — trajectives.
Ce dernier point ne doit pas surprendre : toute la part de créativité, mais encore d’imaginaire, mais enfin d’interprétation nécessite cette souplesse et cette acuité que seul l’œil et le cerveau humains sont en mesure de fournir. La dimension cognitive en œuvre dans l’élaboration du syntaxon et du synsystème relevant en grande partie de la « sphère langagière » du chercheur, n’étant à l’heure actuelle (et probablement encore pour longtemps) que partiellement développée par les machines (quelqu’un décide de l’allocation d’un syntaxon et de son rang à une espèce).
Ainsi, observation, inventaire, tri et organisation des données, analyse des données, aboutissent d’une part à une opération d’interprétation de l’état des lieux cénologiques, d’autre part à une opération que je qualifierai de scénarisation qui permette d’appréhender la dynamique biologique en cours, les interactions entre les différentes cénoses, ou entre les espèces au sein des cénoses. Et, par conséquent, si l’on travaille à l’échelle d’un site naturel, par exemple, la cénotique apporte un éclairage robuste sur les enjeux comme sur les effets d’éventuelles interventions humaines ; seulement alors s’ouvre un nouveau chapitre, celui de la gestion.
Toutes ces étapes me semblent d’égale importance. Celles-ci s’étalent d’ailleurs dans le temps et cette donnée devrait également être prise en compte, on ne peut que difficilement connaître et comprendre les éléments d’une cénose et encore moins un site naturel en quelques heures de passages, le site fut-il peu patrimonial, ou ‘simple’, et les passages échelonnés dans les saisons.
Durant la phase de terrain, le chercheur réalise des relevés eux-mêmes formalisés (aire ou volume minimaux, données physico-chimiques complémentaires, liste des espèces, coefficients d’abondance). Chaque cénologue définira au préalable l’aire minimale, ainsi que l’unité de comptage préférentielle pour le clade dans lequel il opère.
La phase de synthèse consiste à traiter les relevés de manière à rendre évidents les groupes cénologiques ; ceux-ci sont alors comparés à ceux déjà publiés et « typifiés » ou, le cas échéant, présentés nouvellement. Les syntaxons ainsi créés ou nommés sont ensuite insérés par le chercheur, ou après-coup par la communauté scientifique, dans le synsystème.
À part le plaisir de l’inventaire/l’invention, et celui de l’imaginaire, consistant à inventer des formes particulières (les syntaxons), la cénologie présente d’évidentes applications :
- l’application la plus simple, est de considérer le syntaxon, comme la cénose, comme une abstraction, un artefact humain, mais si on acceptait cette dimension subjective au sens anthropique du terme, je pose qu’on pourrait beaucoup plus facilement, non seulement mesurer la rareté et la beauté de certains groupements, ou bien la relative anthropisation de tel ou tel groupement ;
- concernant la connaissance : si l’on pouvait dénommer des « formations animales » comme on est en mesure de nommer des formations végétales (le bas-marais alcalin ; la hêtraie calcicole, par exemple), on gagnerait un temps précieux ; mais surtout on proposerait un début d’intégration, allais-je dire totale, au niveau du biotope ou de l’écosystème ;
- concernant la préservation : c’est l’aspect le plus évident : si l’on pouvait définir par exemple que la cénose des « gastéropodes millimétriques hygrophiles de bas-marais alcalins calcicoles » était protégée (même si les espèces caractéristiques étaient absentes), on pourrait rapidement et facilement protéger de nombreux espaces naturels.
Militer pour le retour de la cénologie, c’est non seulement militer pour l’acceptation de la subjectivité y compris dans les sciences, mais c’est également militer pour une réappropriation par les naturalistes des concepts et méthodes des sciences naturelles. La phytosociologie est un continent isolé du reste des études écologiques, alors qu’elle est l’un de leurs piliers. Gageons que le retour de la biocénologie, diffusé notamment au monde animal, et pourquoi pas bactériologique, viennent enrichir ces études, autant sur le plan fonctionnel, que gestionnaire. Si notre recherche est celle des relations d’intégration, alors nous sommes bien obligés de replacer celle-ci au cœur de celles-là.
L’Ecobiocénotique résulte de la confluence de deux grands courants scientifiques […] Elle représente l’aboutissement des sciences naturelles dites “de terrain” […] elle ne saurait être dépassée un jour par une discipline plus vaste. Elle constitue une limite vers laquelle tendent asymptotiquement, volontairement ou non, tous les efforts des naturalistes qui se vouent à l’étude du milieu[20]Roger Molinier & Pierre Vignes, op.cit..
[Cap au seuil|Sommaire rapide] Avant propos | espace | la vie | monde | les écologies | cénologie | sciences naturelles | interlude | protection de la nature | arbres | anthropocène | catastrophe | herbier/coquillier | ouverture : régions naturelles
Les deux (ou trois) écologies < Pour la cénologie > Sciences naturelles = sciences humaines
References
| ↑1 | Parts de ce texte ont paru sous le titre « What has become of our cenosis? » dans Plant Sociology n°58/2, 2021, et d’autres sous le titre « Éléments pour la description des cénoses animales » dans Folia conchyolologica n°64, 2022. |
|---|---|
| ↑2 | C. Flahault , C. Schröter , Rapport sur la nomenclature phytogéographique, in IIIe Congrès international de botanique, Wildemann, Bruxelles, 1910, Actes du IIIe Congrès international de botanique, vol. 1 , 1910. |
| ↑3 | Josias Braun et Ernst Furrer, « Remarques sur l’étude des groupements de plantes », in Bulletin de la Société Languedocienne de Géographie n°36, 1913, 20-41. |
| ↑4 | Pour ces deux domaines de recherche, toutefois, il faut préciser leur singularité biologique ou leur originalité épistémologique. En effet, dans les deux cas, on constate une très forte interaction avec d’autres groupes d’organismes, les bactéries avec les champignons et les phages, les champignons avec les bactéries et les végétaux, sans parler des relations habituelles (qui nous sont plus familières) que virus, bactéries et champignons entretiennent avec l’ensemble du vivants. Dans le cas des bactéries, des associations de type écologique existent, ce qu’on pourrait appeler avec Charlotte Brives, la pluribiose — voir Charlotte Brives, Pluribiose, Quae, 2024 ; concernant l’association champignon/végétal (+bactérie), communément appelées mycorhizes, les recherches sont littéralement en plein boum, et nos connaissances progressent à grand pas — voir Jean Garbaye, La symbiose mycorhizienne. Une association entre les plantes et les champignons, Quae, 2013. Les deux ouvrages, synthétiques, présentent une roborative bibliographie sur leurs domaines respectifs. On touche alors à la thématique de la symbiose, interaction biologique voisine du mutualisme, du commensalisme, du neutralisme, de l’amensalisme, du parasitisme, de la prédation, autant de relations interspécifiques à caractère trophique. |
| ↑5 | Actes du colloque sur l’écologie de 1950 à Paris, in Colloques Internationaux du Centre National de la Recherche Scientifique, n°XXXIII, Paris, C.N.R.S [Note de Pascal Acot : ‘Les Actes du colloque sont constitués par 4 fascicules de L’Année Biologique (3e série, t. 27, fasc. 2, 4, 6, 7, 1951). Le fascicule 5 étant sauté (pages 161-242), la pagination des fascicules [pp. 76-582] ne correspond pas à la pagination réelle des Actes (350 pages). L’éditeur a donc établi une pagination double. Les références qui suivent renvoient à la pagination réelle et non à celle des fascicules. Les comptes-rendus des discussions sont inclus dans la pagination des communications données en référence.’] ; et voir donc le même Pascal Acot, « Le Colloque international du CNRS sur l’écologie (Paris, 20-25 février 1950) » in Jean Gayon et Claude Debru (éds.), Les Sciences biologiques et médicales en France, 1920-1950, Paris, CNRS-Editions, 1994, p.233-240. |
| ↑6 | Pierre Quézel et Paul Verdier, « Les méthodes de la phytosociologie sont-elles applicables a l’étude des groupements animaux ? Quelques associations ripicoles de carabiques dans le Midi de la France et leurs rapports avec les groupements végétaux correspondants » in Vegetatio Acta Geobotanica n°4, p.165-181. |
| ↑7 | On pourrait citer Rabeler, Künhelt, Strenzke, Sacchi, Schwenke et Quézel & Verdier, Amiet, Mouthon, Frank, etc. tout ceci largement concentré dans les années 50. |
| ↑8 | Roger Molinier, « L’étude des biocénoses marines du Cap Corse », in Vegetatio n°9, p.121–192 & 217–312, 1960 ; Charles-François Boudouresque, « Recherches sur les concepts de biocénose et de continuum au niveau des peuplements benthiques sciaphiles », Vie et Milieu n°21, p.103-136. |
| ↑9 | Jean-Marie Pérès & Jean Picard, « Nouveau manuel de bionomie benthique de la mer Méditerranée », Recueil des Travaux de la Station Maritime d’Endoume n°47(31), p.1-137. |
| ↑10 | Marcel Huet, « Aperçu des relations entre la pente et les populations piscicoles des eaux courantes », in Schweizerische Zeitschrift für Hydrologie n°11, p.332–351, 1949 ; Jean Illiès & Lazăr Botoşăneanu 1963. Problèmes et méthodes de la classification et de la zonation écologique des eaux courantes, considérées surtout du point de vue faunistique, in SIL Communications n°12(1), p.1-57, 1963 ; Jean Verneaux, « Fondements biologiques et écologiques de la qualité des eaux continentales. Principales méthodes biologiques », in P. Pesson et al. (éd.)., La pollution des eaux continentales, Bordas, p.289-345, 1989. |
| ↑11 | Robert Whittaker, « Classification of Natural Communities » in The Botanical Review, n°28/1 : pour les cénoses animales, voir p.62-69 ; dans la bibliographie tous les auteurs cités jusqu’en 1962 sont présents. |
| ↑12 | Reinhard Tüxen (éd.), Biosoziologie, La Hague, Junk, 1966. |
| ↑13 | Roger Molinier & Pierre Vigne, Ecologie et biocénotique, Delachaux & Niestlé, 1971. |
| ↑14 | En réalité, c’est plus compliqué, comme d’habitude. Rioux distingue, de son côté, trois types de groupements : la cénose, la société et la foule. La foule est un groupement sans autre lien que la ressource : ce serait donc un synonyme de la guilde. La société est un groupe avec des liens forts, phylogéniques (familiaux ou spécifiques) s’érigeant en ensemble autonome (la « cité » ), ce qui l’associe à la population : De quelques concepts en biocénotique, op.cit. |
| ↑15 | Jeffrey A. Lockwood, « The Ontology of Biological Groups : Do Grasshoppers Form Assemblages, Communities, Guilds, Populations, or Something Else ? » in Psyche : A Journal of Entomology, en ligne : ttps ://www.hindawi.com/journals/psyche/2011/501983/ |
| ↑16 | Mauritz Franz Mörzer Bruyns, On biotic communities [1947], Station Internationale Géobotanique Méditerranéenne et Alpine, 1950. |
| ↑17 | Gisin (1964) se méfiait des diagrammes en treillis qui selon lui étaient des non-sens ; Gounot lui-même (1961) mettait en garde à l’absence de recul devant les équations ; Guinochet (1968) : « Beaucoup d’écologistes auraient besoin qu’on leur rappelle qu’il faut savoir adapter la précision des mesures à l’objet mesuré ». Citant ces derniers, Boudouresque (1971) surenchérit : « Les chiffres en eux-mêmes ne signifient rien : ils ne valent que par leur contexte, par le sens et les limites qu’on veut bien leur attribuer. Trop de bionomistes réduisent la Statistique à quelques formules qu’ils utilisent à contre sens » . |
| ↑18 | Helmut Gams, « Prinzipienfragen der Vegetationsforschung. Ein Beitrag zur Begriffsklärung und Methodik der Biocoenologie » , loc.cit. |
| ↑19 | Gams, loc.cit., p.298 ; Rioux, op.cit., p.129 ; Mörzer Bruyns, op.cit., p.1-3. |
| ↑20 | Roger Molinier & Pierre Vignes, op.cit.. |